History of Spatial Transcriptomics: Spatial Transcriptomics of Tissue Regions
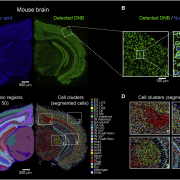
To overcome the limitation of only being able to study gene expression in areas of a tissue that were isolated by LCM, a group at The Spatial Research Lab at the Science for Life Laboratory developed a novel slide-based transcriptomics approach. This group was led by Drs. Joakim Lundeberg, Stefania Giacomello, and Patrik Ståhl (14). The spatial transcriptomics (ST) technology they developed utilized single-stranded DNA probes, spotted on a slide in a manner similar to microarrays. These slide-bound DNA oligo probes contain a spatial barcode to map back to its position on the slide for spatial expression analysis. The oligo also contains a Unique Molecular Identifier (UMI) to differentiate the probes on the same spot. At the 3’ end of the oligo probe, it contains a poly-T tail to capture polyadenylated mRNA released from the permeabilized tissue. The captured mRNA is reverse transcribed resulting in a spatially barcoded cDNA that is prepared as a library for next generation sequencing (NGS) (14). The end result is whole transcriptome sequencing data that can be mapped back to its original location in the tissue. Initially, the ST microarray consisted of ~1000 spots, each with a diameter of 100um. At the end of 2019, the ST technology was acquired by 10X Genomics and commercialized under the name Visium. 10X Genomics increased the number of spots on the array to ~5000 and reduced the diameter of each spot to 55um. At this resolution, each spot covers 1-10 cells. Although this greatly improved the resolution for ST, it is still not able reach the level of single cell or the ultimate goal of subcellular resolution.
In 2019, NanoString Technologies also launched its first spatial transcriptomics platform, GeoMx DSP (Digital Spatial Profiler). This technology enabled multiplexed spatial profiling of not only RNA but also proteins. Unlike Visium’s method of capturing and sequencing RNA via an oligo dT probe, the GeoMx DSP probe uses a unique complementary sequence to directly hybridize to a transcript of interest. This target-specific capture probe is attached to a DSP barcode via a photocleavable linker. The DSP barcode is used to backtrack the location of the transcript in the tissue. These barcodes can be analyzed downstream in the workflow using NanoString’s “n-counter” system or by NGS, depending on the number of genes desired for analysis (~1000 or ~18,000, respectively). Two of the most significant limitations of the GeoMx DSP system are its resolution and single molecule detection approach. Like Visium, GeoMx cannot detect genes expressed in individual cells (resolution can only go down to 100µm).
Top questions about spatial transcriptomics
What is a Unique Molecular Identifier (UMI) for Stereo-seq?
Unique Molecular Identifiers or UMI are short sequences or molecular “tags” contained in each oligo on a Stereo-seq chip. Each oligo within a spot on the Stereo-seq chip has its own UMI to identify that individual oligo.
What are limitations of the Visium and GeoMx Digital Spatial Profiler technologies?
Two of the most significant limitations of the GeoMx DSP system are its resolution and high cost. Both 10x Genomic’s Visium and NanoString’s GeoMx DSP cannot detect gene expression at the single cell level as each spot typically overlaps 5 to 10 cells. The costs to run each of these platforms, including sequencing, is often over $10,000 for each sample.